© Universität Bielefeld
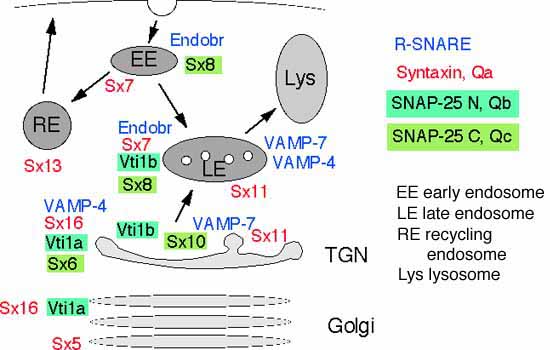
Mammalian SNAREs
Human vti1b can replace yeast Vti1p in certain membrane transport steps if expressed in yeast, indicating that it may have similar functions in mammalian membrane traffic [details]. We are currently studying the role of Vti1a and Vti1b in mammalian membrane traffic, their localization and their SNARE partners.
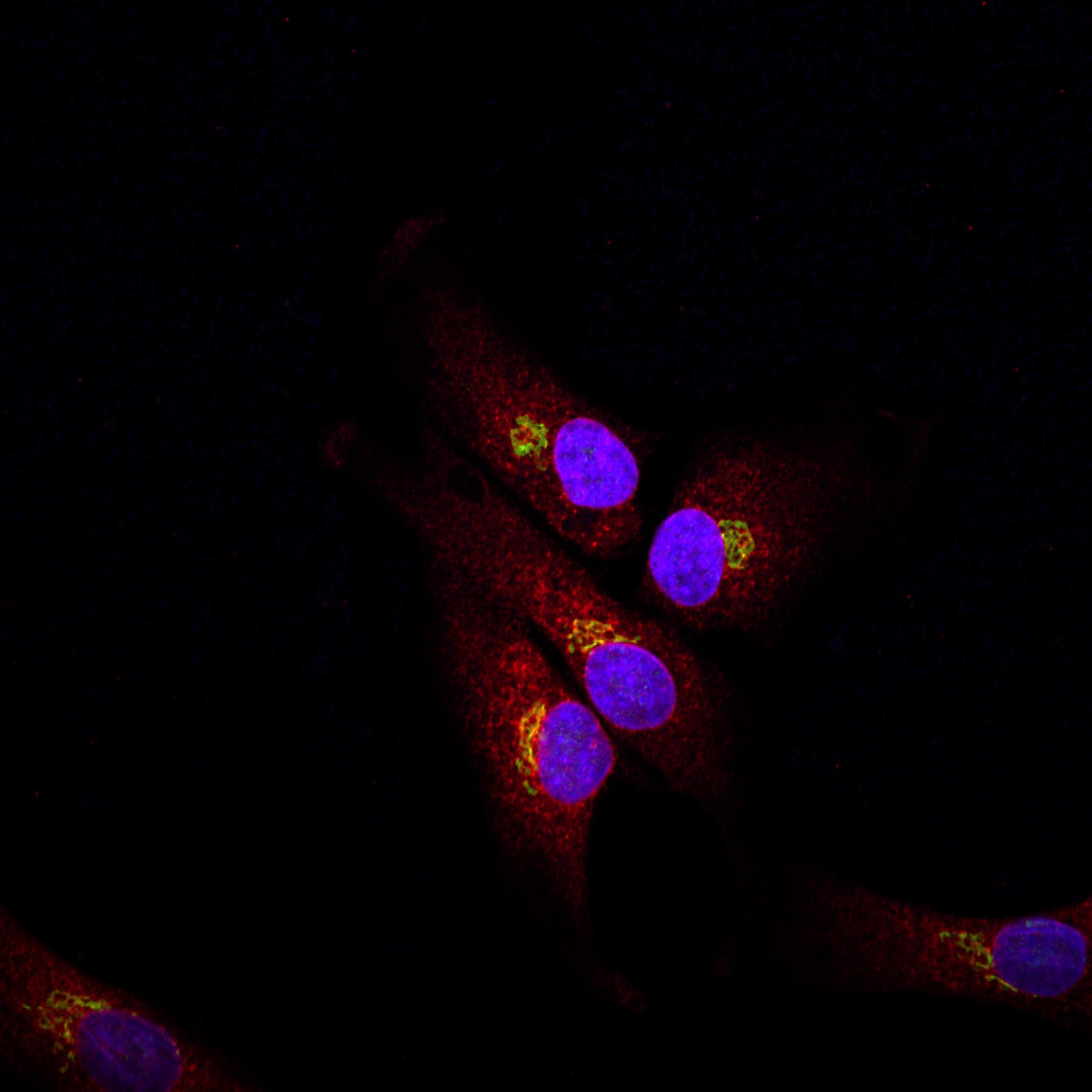
Vti1a and vti1b have a distinct but overlapping localization. Vti1a is found predominantly on the Golgi and the TGN, vti1b mostly on tubules and vesicles in the TGN area and on endosomes [details].
Localization of SNAREs in mammalian cells
We identified the brain specific splice variant vti1a-b which is localized to synaptic vesicles throughout their life cycle [details]. Our data suggest that vti1a-b is not involved in exocytosis but in a separate SNARE complex with VAMP-4, syntaxin 16 and syntaxin 6 [details]. This complex probably functions in a membrane fusion step during recycling or biogenesis of synaptic vesicles. It may be required for fusion with early endosomes during recycling because antibodies against vti1a inhibit fusion of early endosomes in an in vitro assay [details]. Vti1a has also been implicated in intra Golgi traffic (Xu et al. 1998 J. Biol. Chem. 273, 21783).
Vti1b forms a SNARE complex with syntaxin 7, syntaxin 8 and the R-SNARE endobrevin which shares common structural features with the neuronal SNARE complex. Antibodies against any of the components inhibit fusion of late endosomes in vitro and block lysosomal degradation of EGF in vivo [details].
Common structure of SNARE complexes
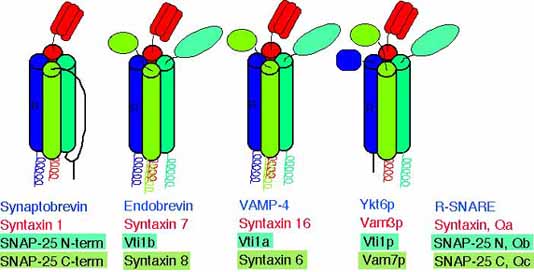
Our data indicate that the SNARE complexes containing the different Vti1 proteins and three additional SNAREs have a similar structure as the complex consisting of synaptobrevin, syntaxin 1 and SNAP-25 despite little similarities in the amino acid sequences.
The N-terminus of vti1b but not vti1a interacts with the ENTH domain of enthoprotin/CLINT/epsinR indicating that this interaction between a SNARE and an ENTH domain is conserved between mammals and yeast [details]. Both, epsinR and Ent3p are involved in the formation of transport vesicles traveling between the TGN and endosomes. EpsinR acts as a cargo adaptor for sorting of vti1b but not vti1a into budding clathrin coated vesicles because knock down of epsinR reduced the amount of vti1b found in clathrin coated vesicles (details). In addition, epsinR shows weak interactions with two other SNAREs, syntaxin 7 und syntaxin 8, which are involved in traffic to the late endosome and to the lysosome together with vti1b [details].


We generated knock out mice deficient for vti1b to obtain an animal model for analyzes of defects in transport to the lysosome in the whole animal and in specific tissues. Vti1b deficient mice had reduced amounts of syntaxin 8 protein while the amounts of syntaxin 7 and endobrevin did not change. These data indicate that vti1b was specifically required for the stability of a single SNARE partner. Most vti1b deficient mice were indistinguishable from wild type mice. About 20% of the vti1b deficient mice were smaller and accumulated multivesicular bodies and autophagosomes in hepatocytes [details]. In cytotoxic T cells absence of vti1b resulted in an impaired release of lytic granules, which belong to the group of secretory lysosomes. A similar phenotype was observed in mice deficient for Vamp8 [details].
In addition, we generated knock out mice for the early endosomal vti1a. These mice are viable without overt phenotypes [details] but the biogenesis of dense-core vesicles in chromaffin cells is impaired [details]. In the absence of vti1a and vti1b mouse embryos die at birth with massive degeneration of peripheral ganglia and impaired neurite outgrowth and axonal guidance. These data suggest that special demands for endosomal membrane traffic could not be met in Vti1a-/- Vti1b-/- neurons. However basic endosomal traffic was functional in Vti1a-/- Vti1b-/- fibroblasts suggesting a surprising compensation by other unrelated SNAREs [details].