Yeast SNAREs

Yeast SNAREs
Baker's yeast (Saccharomyces cerevisiae) is used as a model organism because basic cellular functions are very similar in yeast and humans. For technical reasons it is easier and faster to study yeast as a combination of genetic and biochemical approaches can be used. Results obtained in yeast can be applied to mammals in subsequent projects.
We are interested in the role of SNARE proteins in yeast membrane traffic. We identified a SNARE with low homologies to other SNAREs which functions in retrograde traffic from the Golgi to the ER. It localized to the ER and was therefore named Use1p (unconventional SNARE in the ER, details). The SNARE motifs of Use1p and Sec20p are so divergent that they can't be assigned to either Qb- or Qc-SNAREs.
SNAREs in yeast membrane traffic
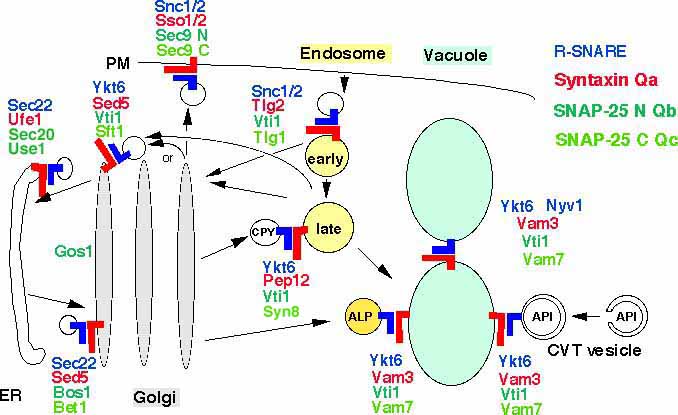
The Yeast Q-SNARE Vti1p and the R-SNARE Ykt6p interact with three different syntaxin related Q-SNAREs in several transport pathways
We concentrate on the Qb-SNARE Vti1p and the R-SNARE Ykt6p. SNAREs were thought to determine specificity in vesicular transport. However, Vti1p interacts functionally with three different t-SNAREs in at least four different transport steps. Vti1p functions with the endosomal t-SNAREs Pep12p in traffic from the trans Golgi network to an endosomal compartment and with the cis-Golgi t-SNARE Sed5p in retrograde traffic to the cis-Golgi (details). Vti1p is also required for three different biosynthetic trafficking pathways to the vacuole, the equivalent of the mammalian lysosome and for homotypic vacuolar fusion. Vti1p interacts with the vacuolar t-SNAREs Vam3p and Vam7p in all vacuolar trafficking steps (details). Ykt6p is the R-SNARE in these three SNARE complexes (details). The ability of Vti1p and Ykt6p to mediate multiple fusion steps requires additional proteins to ensure specificity in membrane traffic. The mutations vti1-Q158R and ykt6-R165Q were introduced into the 0 layers to study its role in SNARE complexes. Cells expressing only ykt6Q were not viable. vti1R cells had severe defects in several transport steps. These data indicate that the arginine in the 0 layer of Ykt6p is essential and that an additional arginine in the 0 layer interferes with function (details).
We are currently investigating the role of the N-terminus of Vti1p in yeast membrane traffic. We identified Ent3p as the first protein interacting with this domain (details). Ent3p belongs to the family of ENTH domain proteins which function in the formation of clathrin coated vesicles. Ent3p also binds to the SNAREs Pep12p and Syn8p which are required for transport from the TGN to the late endosome together with Vti1p and Ykt6p. Pep12p trafficking was altered by removal of Ent3p or inactivation of a sorting signal in Pep12p required for binding to Ent3p. Therefore Ent3p acts as a cargo adaptor for sorting of these SNAREs in TGN to late endosomal traffic (details).


The structure of the complex between Ent3p-ENTH domain and the N-terminal domain of Vti1p was determined. Ent3p interacts via amino acid residues, which are conserved from yeast to man. However, the interaction surfaces are on opposite sides of the three helix bundles in Vti1p and mammalian vti1b (details). Vti1p, Pep12p and Syn8p bind to the same area of Ent3p. However, different amino acid residues were crucial for these interactions (details). The temperature sensitive vti1-3p has two amino acid exchanges in the N-terminal domain, which results in unfolding of this domain. This causes a block in transport to the late endosome and retrograde transport to the trans Golgi network while fusion with the vacuole was possible but slower (details).
We could show that the ENTH domain protein Ent5p acts as a cargo adaptor for the SNARE protein Tlg2p in transport between endosomes and Golgi (details).